
第二节 细胞凋亡的机制
多细胞个体在凋亡过程中大多拥有相似的酶反应过程。死亡受体和线粒体介导的两条通路是经典的细胞凋亡信号传导通路,此外还有近年来研究发现的过度内质网(ER)应激启动的细胞凋亡信号传导通路。
一、死亡受体介导的细胞凋亡
死亡受体属于肿瘤坏死因子(tumor necrosis factor,TNF)基因家族,其共同特征是都有相似的、富含半胱氨酸的细胞外结构域和一个同源的称为死亡结构域(death domain,DD)的胞内序列。死亡结构域一般使死亡受体与胞内凋亡机制相连,但它们有时也会介导与凋亡无关或抑制凋亡的过程。
(一) FasL-Fas系统介导的细胞凋亡
自杀相关因子(factor associated suicide,Fas)是广泛表达于正常细胞和肿瘤细胞膜表面的Ⅰ型受体,其胞质段内由含有60~80个氨基酸组成的死亡结构域及阻抑域(suppresive domain,SD)。Fas的SD被Fas结合磷酸酶-1(fas-associated phosphatase-1,FAP-1)结合后可抑制Fas的诱导凋亡作用。
Fas的配体FasL(Fas ligand)主要表达于T效应淋巴细胞和肿瘤细胞,是呈现于膜表面的Ⅱ型受体。Fas和FasL都是以纯三聚体形式存在,在FasL同靶细胞膜表面的Fas结合后,诱导Fas胞质段内的DD结合Fas结合蛋白(fas associated protein with DD,FADD),FADD再以其N端的死亡效应子域(death effector domain,DED)结合前胱冬肽酶-8或-10,形成一个由Fas-FADD-前胱冬肽酶-8(或-10)3种分子组成的复合体,称为诱导死亡的信号复合体(death inducing signaling complex,DISC),其中前胱冬肽酶-8(或-10)就可以自身催化形成活性的胱冬肽酶-8,由此完成了由Fas介导的死亡信号起始转导(图13-3),接下去进行级联反应激活下游的靶前胱冬肽酶(包括前胱冬肽酶-3,-6,-7)。活性的胱冬肽酶-8尚可催化Bid(Bcl-2族的促凋亡分子),其N端1~60个氨基酸是Bid的抑制区段,N端第60和61位氨基酸之间的肽腱断裂,其活化的C端部分转位到线粒体膜降低其跨膜压,引起线粒体内细胞色素c和前胱冬肽酶-2,-3,-7,-9等死亡因子释放出来。由此通过形成凋亡激活复合体(apoptosome),其中前胱冬肽酶-9自身激活而增大Fas介导的死亡信号转导反应。
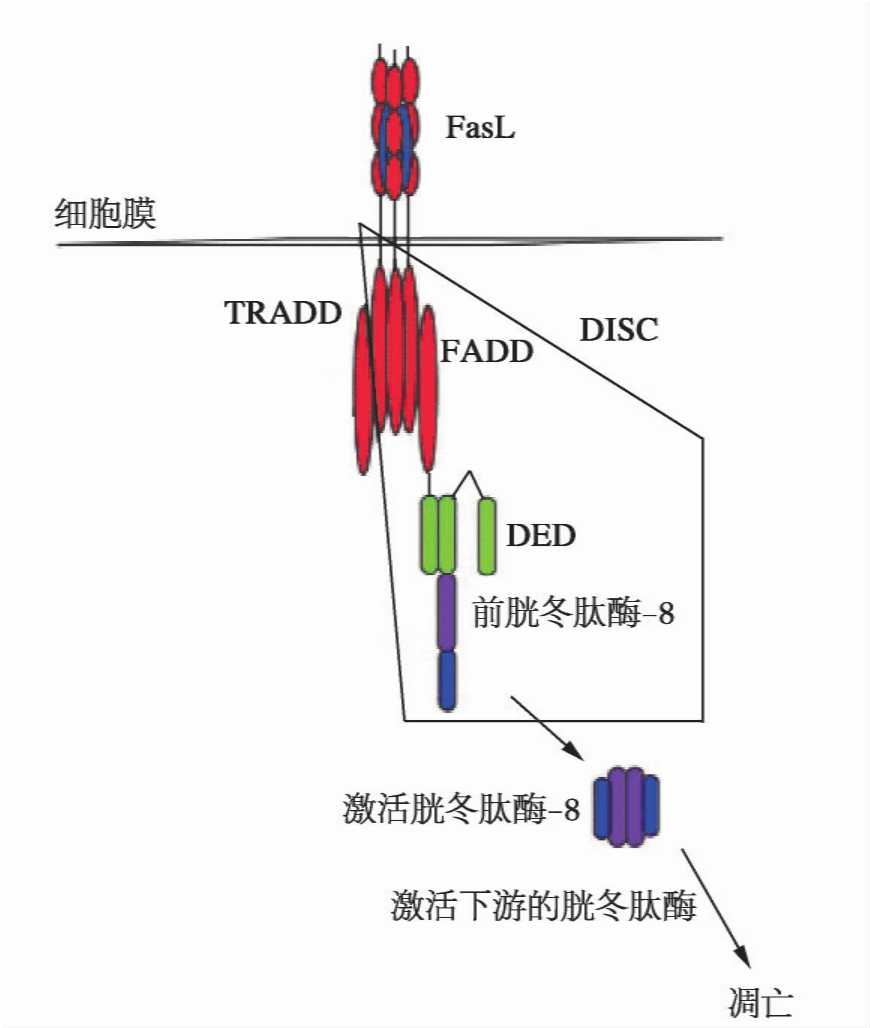
图13-3 FasL-Fas介导的细胞凋亡
(二) TNFα-TNFR1系统介导的细胞凋亡
TNFα及其受体介导的信号通路不是专职负责细胞凋亡的信号转导通路。其受体有TNF-R1和TNF-R2两种亚型,但只有TNF-R1胞质段含有DD,故TNFα引起细胞凋亡的信号转导途径是由TNF-R1介导的。TNFα在诱导细胞凋亡时,与Fas一样,首先自身三聚体化,再与TNFR的胞外区3个分子结合,导致其胞质部分聚集成簇。此后,可通过几条不同的传导通路,诱导细胞凋亡。TNFR既可像Fas一样,直接与FADD结合引发信号下传,也可先与TRADD(TNFR1-associated death domain protein)结合,由于TRADD不含DED结构域,不能直接与Procaspase结合。因此,信号的传递又可通过以下3条通路传导:①TRADD与FADD结合,引发信号下传。②TRADD通过TRAF2(TNF-receptor-associated factor 2)分别导致JNK和NF-κB活化,活化的JNK诱导细胞凋亡,而活化的NF-κB则抑制细胞的凋亡。当JNK和NF-κB同时被激活时,则细胞表现为存活。③TRADD还可选择性地与RIP(receptor interacting protein)结合而激活另一条凋亡通路,RIP与RAIDD(RIP-associated ICH-1/CED-3-homologous protein with a death domain)结合诱导凋亡(图13-4)。
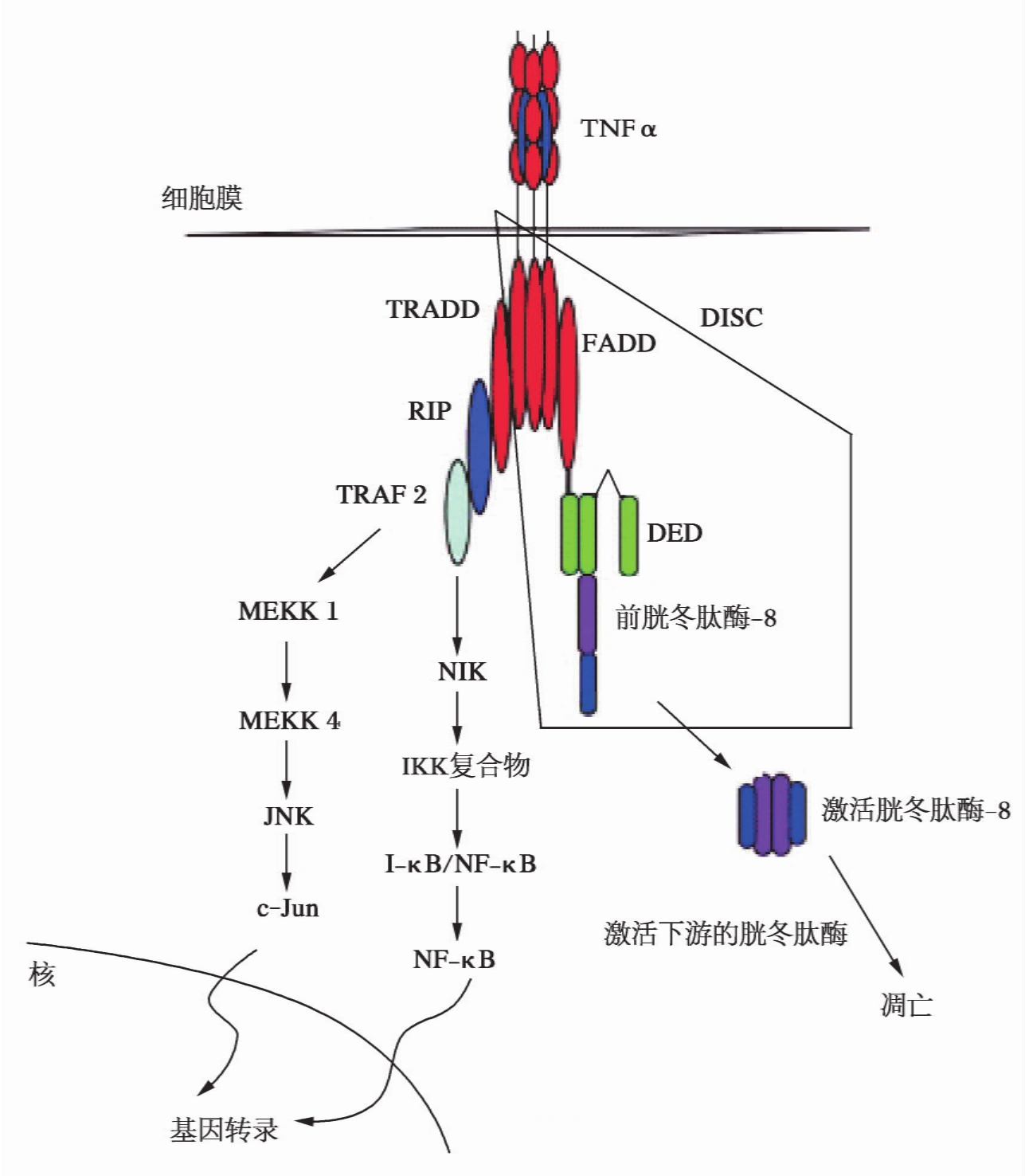
图13-4 TNFα-TNF-R1介导的细胞凋亡
二、线粒体介导的细胞凋亡
在脊椎动物细胞的凋亡过程中,线粒体被认为是处于凋亡调控的中心位置。死亡信号诱使线粒体渗透性转换孔(permeability transition pore,PTP)开启,导致了线粒体跨膜电位(ΔΨm)的崩解,从而使凋亡相关活性物质释放,并继而对胱冬肽酶激活是细胞凋亡实现最根本的生物化学途径,其中Ca2+的释放、质子的渗漏以及细胞色素c的释放等都起到了关键作用。细胞色素c从线粒体膜间隙释放出来后即与胞质中Apaf1(线虫CED-4的同源物)结合活化胱冬肽酶-9,进而激活胱冬肽酶-3,导致细胞凋亡。Bcl-2通过阻止细胞色素c从线粒体的释放来抑制凋亡;而Bax通过与线粒体上的膜通道结合促使细胞色素c的释放而促进凋亡。最近的研究表明,Fas途径中胱冬肽酶-8活化后切割胞质内的Bid(Bcl-2家族成员),使之进入线粒体,引发细胞色素c高效释放。Bid诱导细胞色素c释放的效率远高于Bax。
此外,定位于线粒体上的转录因子p53能与Bak、Bax直接相互作用,这种相互作用解除了Bak与Mcl-1(Bcl-2蛋白家族中的抗凋亡蛋白)的结合,使Bak中的BH3结构域外露,从而处于促凋亡的结构构象;这种相互作用还引起Bak、Bax多聚化,导致线粒体通透性的改变,使各种促凋亡因子释放到细胞质中。
线粒体释放的另一凋亡诱导因子是凋亡诱导因子(apoptosis inducing factor,AIF),其相对分子质量约49605,是一种黄素蛋白。正常生理状态下,AIF被封闭于线粒体中,在细胞发生凋亡时被转运入细胞核。重组的AIF能诱导细胞核中染色质凝集和DNA大规模降解。AIF还能诱导纯化的线粒体释放细胞色素c和胱冬肽酶-9。
线粒体是细胞的能量代谢中心,细胞凋亡过程中对线粒体最直接的损伤是其功能丧失。线粒体功能障碍除导致自由基产生增加、兴奋性氨基酸释放增多、Ca2+超载而引起细胞凋亡外,同时还能直接诱导细胞凋亡。此过程中线粒体的能量贮备起决定性的作用,如果损伤程度相对较轻,ATP保留了提供细胞凋亡所需的能量,则发生细胞凋亡;若损伤严重,ATP急剧耗竭,则发生细胞坏死。有研究表明,细胞内ADP与ATP的比值是细胞存活或凋亡的决定因素,比值低于0.2时细胞存活;比值大于此值细胞则将发生凋亡。
三、ER介导的细胞凋亡
ER是细胞内蛋白质合成的主要场所,也是主要的Ca2+库。ER在维持细胞内Ca2+的稳定以及膜蛋白的合成、修饰和折叠等方面有着关键性作用,近年来研究发现ER在凋亡信号处理过程中也发挥着重要作用。引发ER介导的细胞凋亡这一信号传导通路包括非折叠蛋白反应和Ca2+起始信号等机制。在ER Ca2+平衡的破坏或者ER蛋白过量积累等应激情况下,ER膜上的胱冬肽酶-12基因被诱导表达,同时也导致胞质的胱冬肽酶-7转移到ER表面。胱冬肽酶-7激活胱冬肽酶-12,激活的胱冬肽酶-12裂解胱冬肽酶-3等下游效应蛋白酶,最终导致细胞凋亡。
过度ER应激会出现胞质内Ca2+浓度迅速持续地升高,可以激活胞质中的钙依赖性蛋白酶,又可以作用于线粒体,影响其通透性并导致其膜电位的改变,从而涉及线粒体及细胞色素c的协同作用而使胱冬肽酶激活和细胞凋亡。
四、凋亡相关基因
在细胞凋亡的分子生物学研究过程中,发现了有多种基因及基因家族参与细胞凋亡的调控,其中包括胱冬肽酶基因家族、Bcl-2基因家族、p53基因、Fas/Apo基因及c-myc基因等。
(一)胱冬肽酶基因家族
1993年,人们在C.elegans中发现一个编码细胞死亡的基因ced-3与动物细胞中白细胞介素1β转化酶(interleukin-1β-converting enzyme,ICE)基因结构相似。目前在人类中至少有11种ced-3/ ICE成员被发现。它们都属于半胱氨酸蛋白酶,可以水解底物中特异位点的天门冬氨酸,也可以水解自身特异位点的天门冬氨酸而活化自身,因此,目前称它们为胱冬肽酶半胱氨酸天冬氨酸酶。所有胱冬肽酶均以酶原形式合成,它们可通过自身活化、转位活化或被其他蛋白酶水解活化。
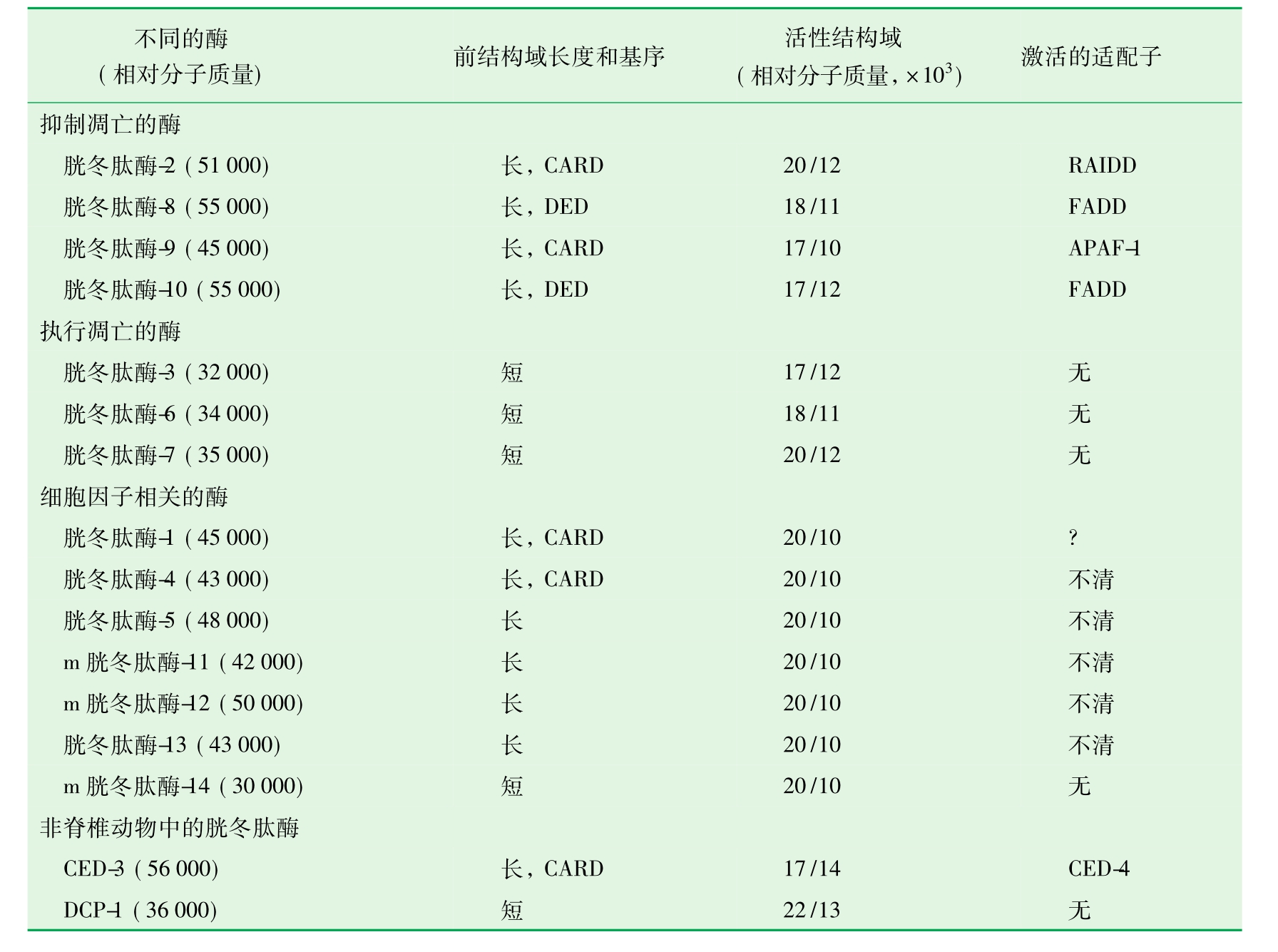
不同胱冬肽酶在结构上有两点显著不同:第一,底物结合位点明显不同,导致不同胱冬肽酶具有各自的底物特异性。第二,不同胱冬肽酶位于N端的前结构域(prodomain)长度和氨基酸组成有很大不同;使胱冬肽酶在凋亡信号转导途径中执行着不同的功能(表13-2)
表13-2 不同胱冬肽酶的特征
(二) Bcl-2基因家族
Bcl-2基因家族包括两大类:一类可抑制细胞凋亡,如Bcl-2、Bcl-xL、Bcl-w、Mcl-1和Bfl-1/A1等;另一类可促进细胞凋亡,如Bax、Bak、Bok、Bad、Bid、Bik、Bim、Hrk/dp5和Noxa等。
许多Bcl-2家族成员的C端都含有疏水尾巴将蛋白质定位在线粒体外膜、ER和核被膜上。抑制凋亡的Bcl-2成员都含有Bcl-2同源区(BH)BH1、BH2、BH3、BH4,其中BH1、BH2和BH3区在一级结构上相距较远,但在三级结构上它们位于同一位置,并形成一个疏水裂缝,与促凋亡蛋白的BH3区结合。
Bcl-2家族成员中抑制凋亡的蛋白与促进凋亡的蛋白之间可形成异二聚体,调节细胞的凋亡,还可通过磷酸化和脱磷酸化修饰其活性。另外,有些成员与线粒体膜上小孔的形成有关,可改变线粒体膜的通透性。位于线粒体外膜上的电压依赖阳离子通道(voltage-dependent anion channel,VDAC)是Bcl-2家族成员作用的靶目标之一,如Bcl-2和Bcl-XL可关闭此通道,而Bax和Bak可使它开放。因此,Bcl-2家族也可通过改变线粒体的通透性调节细胞的凋亡。
(三)Fas和Fas-L
Fas又称为APO-1,属于死亡受体(death receptor)家族,家族中还有TNFR1、TNFR2、CD30、CD40和CD27等,是Ⅰ型膜蛋白,其细胞外部分含2~6个重复的半胱氨酸富集区,成员之间有25%的同源性;相反,胞质部分则很少有同源性。1993年,人白细胞分型国际会议统一命名为CD95。
Fas基因编码产物为相对分子质量45000的跨膜蛋白,分布于胸腺细胞、T细胞、B细胞、NK细胞、内皮细胞、上皮细胞以及皮肤角质形成细胞等。
FasL是TNF家族成员之一,家族中还包括CD30配体、CD40配体、CD27配体、4-1BB配体和TRAIL(TNF-related apoptosis-inducing ligand)等。Fas L以Ⅱ型膜受体的形式合成,即N端位于胞质中。不同成员之间变异较大,C端延伸到细胞外,成员之间有20%~25%的同源性。
Fas与FasL组成Fas系统,两者的结合导致靶细胞走向凋亡。Fas和FasL可因基因突变而丧失功能,从而导致淋巴增殖性疾病发生以及自身免疫加剧,导致组织破坏。
(四)p53基因
p53基因是一种抑癌基因,其生物学功能是在G2期监视DNA的完整性。如有损伤,则抑制细胞增殖,直到DNA修复完成。如果DNA不能被修复,则诱导其凋亡。研究发现丧失p53功能的小鼠胸腺细胞对糖皮质激素诱导的凋亡反应和正常细胞相同,而对辐射诱导的凋亡不敏感。
(五)c-myc基因
c-myc的主要作用是参与转录,而在转录过程中可以激活启动而诱导细胞周期进程和分化,也可以抑制启动而导致阻止细胞分化或程序化死亡,因此它既是激活因子,又是抑制因子。
除了以上凋亡相关基因外,随着研究的不断深入,还发现存在其他多种可能与凋亡调控有关的基因,只是有些研究起步较晚,许多机制尚无法阐明。





